Evoluce
O evoluci obecně, a u včel specielně
Evoluce vysvětluje vznik a vývoj života, obecně, v tzv. evolučních větvích a u druhů
Nejdříve historický přehled

Pozn.:
prokaryotické buňky - jednoduché a malé buňky bez jádra a dalších organel; bakterie a sinice;
eukaryotické buňky - jsou mnohem větší, mají jádro a další buněčné organely; rostlinné a živočišné buňky.
Teorie evoluce
V našem povědomí je zakořeněn darwinistický princip evoluce, tak ho známe z učebnic, médií apod. Charles Darwin ve druhé polovině 19. stol. jako první podal souhrnné vysvětlení vývoje života včetně hybné síly evoluce - tou je přírodní výběr. Ten v důsledku neustálého zápasu uvnitř druhu o zdroje postupně vytváří a modifikuje účelné vlastnosti organismů (vnější podobu, funkce orgánů, modely chování). Byla to skutečná revoluce v dosavadním chápání původu a vývoje života lidstvem.
Darwin přesto uspokojivě nevysvětlil, jak se účelné vlastnosti přenáší z generace na generaci. Ani nemohl, protože už za Darwinova života G. Mendlem objevený (a publikovaný) princip dědičnosti odborný svět našel až počátkem 20. stol. Darwinismus neposkytl ani představu, jak vlastně vznikají nové druhy. To ale vykonali jeho následovníci - neodarwinisté spojením evoluční biologie a klasické i populační genetiky, a rozpracováním teorie speciace (vytváření druhů).
I vývojová teorie se vyvíjí. Dnes je zřejmé, že zdaleka ne jen nesmlouvavý přírodní výběr je zodpovědný za utváření vlastností živých organismů - významnou roli často hraje i náhoda v osudu genů (genetický drift) a další dnes známé procesy. A ještě zásadní věc - darwinistický princip evoluce není platný pro pohlavně se množící (sexuální) druhy. Plynulé přizpůsobování se druhu novým podmínkám působením přírodního výběru funguje jen u druhů asexuálních (např. bakterie) a ještě pak u samosprašných rostlin. U sexuálních druhů Darwinův princip nemůže fungovat, protože genetické založení organismu je v každé generaci jiné v důsledku kombinování genů od rodičů. Výrazně se tedy s každou generací mění vlastnosti potomků, tzn. že vysoká biologická zdatnost jedince, díky níž má reprodukční výhodu před jinými, se v dalších generacích rozplývá. U asexuálních organismů je tomu přesně naopak - genetické založení jedince je skoro v každé generaci beze změn, biologická zdatnost jedince se tak přenáší beze změny z generace na generaci. Díky tomu v populaci postupně převládnou linie lépe přizpůsobené daným podmínkám, ostatní jsou postupně výběrem vytlačeny. Občasnou malou změnu přináší jen mutace (náhodná změna některého genu - alely) a ta může přinést i postupnou změnu vlastnosti v celé populaci, pokud jde o příznivou - výhodnou mutaci. Touto cestou se výhodná mutace v populaci rozšíří - fixuje se.
Jak je tomu tedy u sexuálních druhů? V 60. a 70. letech minulého století přinesl dobré vysvětlení W.D. Hamilton, rozvinul a zpopularizoval je R. Dawkins. Zdůvodnili, že přírodní výběr podle Darwina probíhá i u sexuálních druhů, ale ne na úrovni organismů, nýbrž na úrovni genů. Jednotlivé alely (alela je forma genu) mezi sebou soutěží o to, která bude úspěšnější, která se v populaci více namnoží a přejde do většího počtu organismů. Tento princip je znám jako teorie sobeckého genu, jinak vyjádřeno teorie mezialelické soutěže. Podle ní se jednotlivé alely navenek jeví jako soutěžící jednotky, a organismy jsou jejich pouzí nositelé. Teorie sobeckého genu dovede vysvětlit mnohé evoluční děje u pohlavně se množících druhů, i některé současné projevy organismů, jako např. boj o zdroje, působení přírodní selekce, příbuzenský výběr, altruismus. Nicméně v poslední době se ukazuje, že ani pojetí sobeckého genu nedovede uspokojivě vysvětlit vznik nových druhů, jejich vývoj a další biologické jevy a procesy. Alely v podstatě mezi sebou skutečně soupeří, ale platí to jen v omezené míře, naplno jen v určitých, časově omezených situacích.
Ještě obecnější teorii zveřejnil nedávno český vědec prof. Jaroslav Flegr z Karlovy univerzity. Hlavním nedostatkem genostředného konceptu sobeckého genu totiž je, že převážná většina znaků sexuálních organismů je určena více než jedním genem (polygenní dědičnost) a projev určité alely často závisí na tom, jaké další alely s ní spolupůsobí při utváření znaku. Platí tedy stejná výhrada jako u platnosti Darwinovy teorie individuálního přírodního výběru, tentokrát vztažená na výběr jednotlivých alel. Omezení předchozích teorií dokáže překonat Flegrova teorie zamrzlé evoluce. Ta vysvětluje, že v době existence sexuálního druhu existuje krátké počáteční období, kdy nově vznikající druh tvořený jen malým počtem jedinců je izolovaný od mateřské populace. V důsledku toho se v krátké době úzkým příbuzenským pářením odstraní veškeré genetické rozdíly mezi jedinci této malé populace. Geny jsou v každé generaci ve stejné kombinaci a proto vlastnosti všech jedinců takřka shodné - proto může působit přírodní výběr. Navíc taková nová populace je mateřské populaci podobná co do genetické výbavy jen velmi málo, protože díky své malé početnosti si odnesla jen malou část alel. Tím je dáno, že z oddělené populace může vzniknout nový druh. Ovšem velká většina takto oddělených populací zanikne, protože vzhledem k úzké genetické výbavě nejsou schopné se včas přizpůsobit novým podmínkám. Když takovýchto evolučních pokusů proběhne dost velký počet, některý se podaří a nová populace se cestou selekce a hromadění příznivých mutací rychle promění v nový druh. Popsané období velkých proměn vznikajícího nového druhu a přizpůsobování se podmínkách působením přírodního výběru je období plasticity druhu. V historii druhu trvá přibližně 1-2% doby jeho existence. Možných způsobů vzniku nového druhu, neboli speciací, je několik. Zde nastíněný vznik nového druhu je alopatrická speciace.
Nová populace (vznikající druh) se postupně početně zvětší, obydlí určitý areál, vytvoří si tzv. reprodukční bariéry znemožňující případné splynutí s mateřským druhem, mění svoji genetickou skladbu vlivem intenzivní selekce, genetického driftu, hromadění nových mutací a tak se postupně stává konzervativnějším, až se téměř úplně přestane geneticky měnit - přejde do fáze evolučního zamrznutí. Z genetického hlediska se druh dostává do genetické homeostáze - rovnováhy. V tomto evolučně zamrzlém stavu je druh schopen se měnit cestou selekce jen v malé míře, jež se dá nazvat gumovost. Jsou to změny převážně už ne trvalé, protože pokud dočasný vliv prostředí pomine, celá populace se geneticky vrátí do původního stavu - už není plastická, jen gumová. Pokud však dojde k podstatným změnám prostředí, druh už není schopen se jim přizpůsobit a je odsouzen k zániku. Z některé od něho odštěpené malé populace se může vytvořit nový druh díky své mnohem větší plasticitě, v opačném případě druh zaniká a s ním i celá vývojová větev, jako se stalo např. s Neandrtálci. Ovšem každý druh čeká jeho vymření. Doba trvání savčích druhů je přibližně 5 mil. let, trvání mořských plžů a mlžů je asi 10-20 mil. let.
Evoluce včel
Vývoj včely medonosné se dá posuzovat porovnáváním stávajícího stupně vývoje jednotlivých druhů a zvl. nejbližších příbuzných. Podle evolučního biologa J. Maynarda Smithe prošel vývoj živých forem osmi tzv. evolučními přechody, z nichž poslední je vznik sociálních společenství . Když pomineme člověka, tak vysokého stupně vývoje dosáhly také včely. Vysoká organizovanost sociálního druhu je pro některé druhy zřejmě výhodná, ale zase ne natolik, aby v evoluci převládla tato forma. Některé blízko příbuzné druhy včel žijí také sociálně, ale na různém stupni, jiné nadále soliterně - samotářsky. Z druhů hmyzu vytvářejí vysoce eusociální společenství (tzn. vytvářejí trvalá společenství) včelám poměrně vzdálení termiti z rodu Isoptera. Z řádu Hymenoptera (blanokřídlí) potom do čeledi Formicidae patří vysoce eusociální mravenci a nejblíže včele medonosné z blanokřídlých jsou vosy a sršni z nadčeledi Vespoidea, žijící primitivně eusociálně (tzn. zimu přežívá jen matka, ta společenství zakládá každý rok znovu). Ještě blíže včelám medonosným mají čmeláci ze společné čeledi Apidae (včelovití), kteří ale žijí primitivně sociálně (s výjimkou soliterních parazitických pačmeláků) a ještě blíže z podčeledi Apinae jsou to bezžihadlé tropické druhy včel rodu Mellipona a Trigona, jež tvoří trvalá společenství - viz snímek:

Vedle čeledi Apidae je známo hned několik dalších čeledí včel, jež jsou na různém stupni vývoje z hlediska stupně sociálního života, od solitérních druhů až po primitivně eusociální druhy v čeledi Halictidae (ploskočelkovití), z rodu Lasioglosum. Poznání rozdílů v jejich bionomii je z hlediska evoluce velmi zajímavé. Příbuznost uvedených taxonů hmyzu je jasnější z tabulky, kde je zároveň barevně zaznačeno, ve kterém taxonu se nacházejí primitivně a vysoce eusociální druhy.
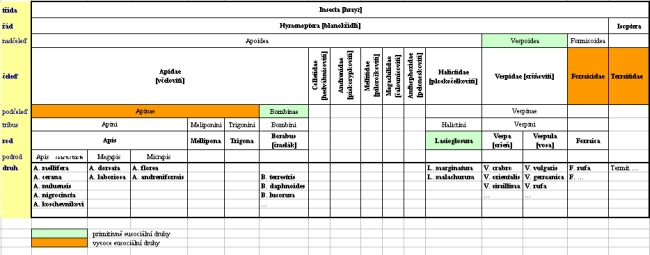
V již zmíněné čeledi Apidae je hospodářsky významný rod Apis s dosud známými 9 druhy společensky žijících včel vysoce eusociálních. Čtyři z těchto druhů staví volně v přírodě na větvi jediný plást (příklad na obr. - plást včely květné, Apis florea), nemají vyvinutou komunikaci tak jako ostatní druhy včel téhož rodu - při vykonávání sdělovacích tanečků musí vidět oblohu.

Zbývajících pět druhů včel staví svá hnízda tvořená několika plásty převážně v dutinách, čtyři z nich také žijí v Asii. Poslední, pro nás nejvýznamnější druh Apis mellifera - včela medonosná, je domovem v Africe, Evropě a jen částečně v Asii, v její západní části. Tvoří poměrně velká společenství, narozdíl od asijských druhů migruje jen výjimečně. Včela medonosná vytvořila řadu geografických plemen tak jak se přizpůsobila různým podmínkách své domoviny.
Vysoce eusociální způsob života je umožněn řadou vlastností, které se u jednotlivců musely postupně vyvinout - anatomické a fyziologické znaky, schopnosti dorozumívání, společně zajišťovaná stavba díla, udržování hygieny hnízda, termoregulace, tvorba zásob potravy, vytvoření kast (matka, trubci, dělnice), utlumení sexuality většiny členů společenství - vznik sterilních dělnic, a s tím související tvorba specialistů na určité činnosti (dělba práce), zvláštnosti rozmnožování, atd. Solitérní druhy většinu uvedeného nemají zapotřebí. Sociální společenství, tedy i včelstva, zřejmě poskytují druhu takové výhody, jež vyváží cenu spojenou se zvládnutím všech jejich dovedností a tělesných i fyziologických předpokladů v průběhu evoluce. Někteří badatelé (Moritz, Southwick, 1992; Moritz, Fuchs, 1998) zavedli pro včelstvo pojem “superorganismus”, když jeho jednotlivé složky v některých anatomických a funkčních ohledech připomínají orgány či buňky vyšších organismů včetně způsobů jejich řízení (např. specifické žlázy a feromony u včel ve funkci hormonů u savců). Ačkoliv pojem superorganismus nevystihuje přesně status včelstva, v praktickém chovu včel je výhodný, protože včelstvo z hlediska ošetřování i šlechtění lze považovat za jakoby jeden ucelený organismus. Ovšemže je to jen určité zjednodušení, vždyť jedinci společenství (nejčastěji včely - létavky) často dočasně odcházejí mimo společenství - odlétají a vracejí se zpět, vně jsou schopni nějaký čas i samostatně žít.
Evolucí sociálních forem, vztahy uvnitř a jejich genetickou skladbou a fungováním se zabývala a zabývá řada výzkumníků a vědců. Zdaleka ne vše je objasněno. Např. otázky tzv. altruismu v hmyzích společenstvech nejsou ve všech souvislostech jasné. Altruismus je jednání jednotlivců ve prospěch celku, neboli sebeobětování. Ukazuje se, že v hmyzích společenstvech je altruismus zcela podřízen potřebám celku, narozdíl např. od dočasných společenstev vyšších obratlovců (stáda zvířat, hejna ptáků, ryb, aj.) i jiných členovců (např. kolonie pavouků, samotářských včel, aj.), kde altruismus jednotlivců přináší prospěch všem i každému jednotlivci a tento prospěch jednotlivec využívá skrze biologickou zdatnost i pro svoji reprodukci. U včel se naproti tomu projevuje absolutní altruismus - včely dělnice konají vše ve prospěch celého společenství, neprojevují žádnou sobeckost ani příbuzenský výběr (nepotismus), jak dokládají některé výzkumy (např. Tarpy aj., 2004; Moritz aj., 2005). Pokud by se dělnice ve včelstvu chovaly sobecky, potom by se jejich geny, sdílené s pohlavními kastami, nemusely přenést do další generace, pokud by uhynulo celé včelstvo, příp. by se snížila jejich šance na přenos, pokud by klesla biologická zdatnost celého včelstva. Sobeckost dělnic a nepotismus by působil negativně. Přínos plynoucí z dělničího altruismu se tudíž má šanci uplatnit jen cestou přenosu přes pohlavní kasty - matky a trubce. Individuální výběr se uplatňuje na úrovni včelstev, uvnitř mezi členy včelstva má podřadnou úlohu nebo žádnou.
Mimochodem - tak se dá vysvětlit, proč se Dawkinsovi v jeho knize “Sobecký gen” (český překlad z r. 1998, str.164) nepodařilo vysvětlit mechanismus působení sobeckých genů ve společenství včel, když na závěr úvah o blanokřídlých a o včelách přiznává: “Skutečnost není tak uhlazená a uspořádaná. Například vůbec nejznámější společenský hmyz, včela medonosná, dělá, jak se zdá, všechno naprosto “špatně”. … Už mi z toho začíná maličko třeštit hlava, a tak je nejvyšší čas toto téma uzavřít.”.
Se sociálním charakterem včel úzce souvisí genetická skladba včelstva, kde v důsledku polyandrie matek (páření s více trubci) je několik trubčích podrodin s příbuzností v nich 0,75 (supersestry), mezi podrodinami je příbuznost 0,25 (polosestry). Polyandrie tedy zvyšuje polymorfismus uvnitř včelstva, tak mu dává poměrně široký arzenál geneticky podmíněných schopností reagovat na různé situace - jde o tzv. sociální heterozi. Pozoruhodný je i způsob rozmnožování, který probíhá na dvou úrovních - nepohlavní množení, neboli rojení, zabezpečuje udržování hustoty druhu v prostoru a zároveň šíření genů matky, pohlavní množení zabezpečuje přenos genů do příští generace a jejich šíření v prostoru prostřednictvím mladých matek a trubců.
S použitím termínů evoluční biologie lze říci, že včela medonosná (stejně jako další druhy včel, mravenci, termiti) je druhem vysoce komplexním (komplexita = složitost) a zároveň vysoce organizovaným. Přitom lze tvrdit, že stupeň sociality včely medonosné je z těch nejvyšších, protože se vyvinula do takové míry, že je podmínkou trvalé existence druhu.